

Struktur, Jenis, Jenis, Fungsi Intermediate
- 4484
- 1344
- Jessie Harvey
Itu filamen menengah, Juga dikenal dalam literatur sebagai "IFS" (dari bahasa Inggris Filames menengah), adalah keluarga protein berserat sitosolik yang tidak larut yang ada di semua sel eukariotik multiseluler.
Mereka adalah bagian dari sitoskeleton, yang merupakan jaringan filamen intraseluler yang terutama bertanggung jawab atas dukungan struktur sel dan berbagai proses metabolisme dan fisiologis seperti pengangkutan vesikel, pergerakan dan perpindahan sel, dll.
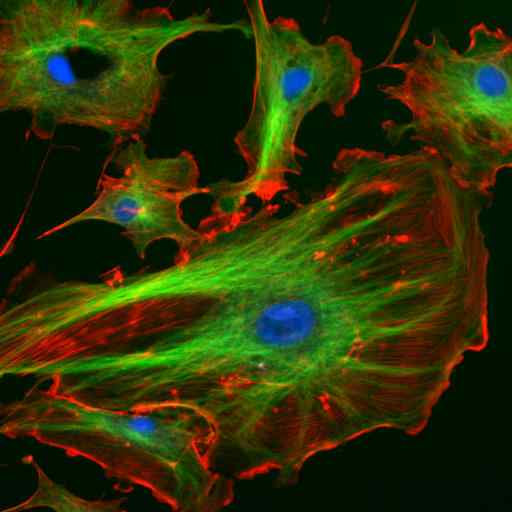
 Mikroskopi dengan imunofluoresensi dua protein filamen perantara astrosit (vimentina dan gfap) (sumber: gerryshaw [cc by-sa 3.0 (https: // createveCommons.Org/lisensi/by-sa/3.0)] via Wikimedia Commons)
Mikroskopi dengan imunofluoresensi dua protein filamen perantara astrosit (vimentina dan gfap) (sumber: gerryshaw [cc by-sa 3.0 (https: // createveCommons.Org/lisensi/by-sa/3.0)] via Wikimedia Commons) Bersama dengan mikrotubulus dan mikrofilamen, filamen menengah berpartisipasi dalam organisasi spasial organel intraseluler, dalam proses endositosis dan eksositosis dan juga dalam proses pembelahan sel dan komunikasi antar seluler.
Filamen perantara pertama yang dipelajari dan dijelaskan adalah keratin, salah satu jenis protein pertama yang strukturnya dianalisis dengan difraksi x -ray pada 1930 -an.
Konsep filamen perantara, bagaimanapun, diperkenalkan pada 1980 -an oleh Lazarides, yang menggambarkan mereka sebagai "integrator mekanik ruang seluler", ditandai dengan kelilin mereka dan kemampuan mereka untuk mereensasi In vitro Setelah denaturasinya.
Banyak penulis dianggap sebagai elemen "bantalan" stres untuk sel hewan, karena mereka adalah filamen yang lebih fleksibel daripada mikrotubulus dan mikrofilamen. Tidak hanya ditemukan di sitoskeleton, tetapi mereka juga bagian dari nucleoesqueleto.
Berbeda dengan komponen berserat lain dari sitoskeleton, filamen menengah tidak berpartisipasi langsung dalam proses mobilitas sel, tetapi bekerja dalam pemeliharaan struktural dan resistensi mekanik sel.
[TOC]
Struktur
Sumber: http: // rsb.Info.Nih.GOV/IJ/Images/[Domain Publik] Filamen menengah memiliki diameter perkiraan 10 nm, karakteristik struktural yang dengannya mereka dinamai, karena ukurannya adalah antara ukuran yang sesuai dengan myosin dan filamen aktin, yang masing -masing sekitar 25 nm dan 7 nm, masing -masing.
Mereka berbeda secara struktural dari dua jenis filamen sitoskeleton lainnya, yang merupakan polimer protein globular, di mana unit konstituennya berbeda protein berserat α-helicoidal dengan panjang besar yang dikelompokkan satu sama lain untuk membentuk struktur dalam bentuk tali atau tali.
Semua protein yang membentuk filamen menengah memiliki organisasi molekuler yang serupa, yang terdiri dari domain α-helicidal atau "tali" yang memiliki jumlah yang berbeda dari segmen "kumparan" dengan ukuran yang sama.
Domain heliks ini diapit oleh "kepala" non-terminal dan "ekor" non-hypoidal di ujung terminal-C, dan keduanya bervariasi dalam urutan dan urutan asam amino.
Dalam urutan kedua ekstrem ini adalah alasan konsensus yang umum untuk 6 jenis filamen perantara yang diketahui.
Pada vertebrata, domain "tali" dari protein filamen sitosolat menengah memiliki sekitar 310 limbah asam amino, sedangkan protein sitosolik dari invertebrata dan laminasi nuklir memiliki sekitar 350 asam amino dengan panjang asam amino dengan panjang 350.
Dapat melayani Anda: Reptil: Apa itu, karakteristik, klasifikasi, reproduksiPerakitan
Filamen menengah adalah struktur "perakitan diri" yang tidak memiliki aktivitas enzimatik, yang juga membedakan mereka dari rekan sitoskeletal mereka (mikrotubulus dan mikrofilamen).
Struktur ini pada awalnya dirakit sebagai tetramer protein filamen yang terdiri dari mereka di bawah pengaruh hanya kation monovalen.
Tetramer ini panjangnya 62 nm dan monomernya dikaitkan satu sama lain secara lateral untuk membentuk "satuan panjang" (UFL, bahasa Inggris Filma lobet unit), Apa yang dikenal sebagai fase 1 dari perakitan, yang terjadi dengan sangat cepat.
UFL adalah prekursor filamen panjang dan, karena dimer yang terdiri dari mereka mengikat satu sama lain dengan cara antiparal dan terhuyung -huyung, unit -unit ini memiliki domain pusat dengan dua domain mengapit melalui fase 2 dari perpanjangan terjadi, di mana penyatuan longitudinal longitudinal UFL lainnya diberikan.
Selama apa yang disebut sebagai fase 3 dari rakitan, pemadatan radial diameter filamen terjadi, yang menghasilkan filamen perantara matang dengan diameter lebih atau kurang 10 nm.
Fungsi
Fungsi filamen menengah sangat bergantung pada jenis sel yang dipertimbangkan dan, dalam kasus hewan (termasuk manusia), ekspresi mereka diatur dengan cara spesifik jaringan, sehingga juga tergantung pada jenis jaringan daripada di bawah di bawah belajar.
Epitel, otot, mesenchymal dan glials dan neuron memiliki berbagai jenis filamen, berspesialisasi sesuai dengan fungsi sel -sel tempat mereka berada.
Di antara fungsi -fungsi ini, yang paling penting adalah pemeliharaan struktural sel dan resistensi terhadap stres mekanik yang berbeda, karena struktur ini memiliki beberapa elastisitas yang memungkinkan mereka untuk membasahi berbagai jenis kekuatan yang dipaksakan pada sel.
Jenis filamen perantara
Protein yang merupakan filamen menengah milik keluarga besar dan heterogen dari protein filamen yang berbeda secara kimiawi tetapi dibedakan dalam enam kelas sesuai dengan homologi urutannya (I, II, III, IV, V dan VI).
Meskipun tidak terlalu umum, berbagai jenis sel, dalam kondisi yang sangat khusus (pengembangan, transformasi sel, pertumbuhan, dll.) Mereka dapat ikut mengekspresikan lebih dari satu kelas protein yang membentuk filamen menengah
Kelas I dan II filamen menengah: asam dan keratin basa
Keratin mewakili sebagian besar protein filamen menengah dan, pada manusia, ini mewakili lebih dari tiga perempat filamen menengah.
Mereka memiliki bobot molekul yang bervariasi antara 40 dan 70 kDa dan berbeda dari protein filamen menengah lainnya karena kandungannya yang tinggi dalam glisin dan serin.
Dapat melayani Anda: landak: karakteristik, habitat, reproduksi, makananMereka dikenal sebagai asam dan keratin basa untuk titik isoelektriknya, yaitu antara 4.9 dan 5.4 untuk keratin asam dan antara 6.1 dan 7.8 untuk dasar -dasar.
Dalam dua kelas ini sekitar 30 protein telah dijelaskan dan hadir terutama dalam sel epitel, di mana kedua jenis protein "co-polimerize" dan membentuk filamen senyawa.
Banyak keratin case dari filamen menengah ditemukan dalam struktur seperti rambut, kuku, tanduk, paku dan cakar, sedangkan kelas II adalah yang paling melimpah dalam sitosol.
Kelas III filamen menengah: protein tipe demine/vimentin
Falls adalah protein asam 53 kDa yang, tergantung pada tingkat fosforilasi, memiliki varian yang berbeda.
Beberapa penulis juga menyebut filamen jatuh sebagai "filamen perantara otot", karena kehadirannya cukup terbatas, meskipun dalam jumlah kecil, semua jenis sel otot.
Dalam myofibrils, jatuh ditemukan di baris z, jadi diperkirakan protein ini berkontribusi pada fungsi kontraktil serat otot saat bekerja di penyatuan myofibrillas dan membran plasma.
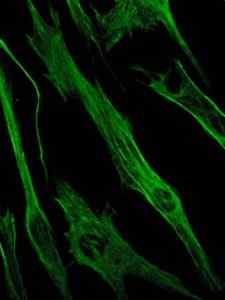 Foto pewarnaan protein vimentin, protein filamen perantara sel epitel dan embrionik (sumber: Viktoriia Kosach [CC BY-SA 4.0 (https: // createveCommons.Org/lisensi/by-sa/4.0)] via Wikimedia Commons)
Foto pewarnaan protein vimentin, protein filamen perantara sel epitel dan embrionik (sumber: Viktoriia Kosach [CC BY-SA 4.0 (https: // createveCommons.Org/lisensi/by-sa/4.0)] via Wikimedia Commons) Pada gilirannya, vimentin adalah protein yang ada dalam sel mesenchymal. Filamen menengah yang dibentuk oleh protein ini fleksibel dan telah ditentukan bahwa mereka menolak banyak perubahan konformasi yang terjadi selama siklus sel.
Ini ditemukan dalam fibroblas, dalam sel otot polos, dalam sel darah putih dan sel -sel lain dari sistem peredaran darah hewan.
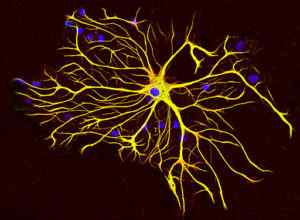
Kelas IV filamen menengah: protein neurofilamen
Juga dikenal sebagai "neurofilamen", filamen menengah semacam ini mencakup salah satu elemen struktural fundamental akson dan dendrit neuron; Mereka sering dikaitkan dengan mikrotubulus yang juga membentuk struktur ini.
Neurofilamen hewan vertebrata telah diisolasi, menentukan bahwa itu adalah triplet protein 200, 150 dan 68 kDa yang berpartisipasi dalam perakitan In vitro.
Mereka berbeda dari filamen menengah lainnya di mana mereka memiliki lengan samping seperti "lampiran" yang diproyeksikan dari pinggiran mereka dan yang bekerja dalam interaksi antara filamen tetangga dan struktur lainnya.
Sel glial menghasilkan jenis khusus filamen menengah yang dikenal sebagai filamen perantara glial, yang secara struktural berbeda dari neurofilamen, karena mereka terdiri dari protein 51 kDa tunggal dan yang memiliki sifat fisikokimia yang berbeda.
Kelas V filamen menengah: filamen lamina nuklir
Semua lembar yang merupakan bagian dari nucleoesqueleto sebenarnya adalah protein filamen menengah. Ini memiliki berat molekul antara 60 dan 75 kDa dan ditemukan di inti semua sel eukariotik.
Dapat melayani Anda: Trichuris trichiuraMereka sangat penting untuk organisasi internal daerah nuklir dan untuk banyak fungsi organel penting ini untuk keberadaan eukariota.
Kelas VI filamen menengah: Nestines
Jenis filamen perantara ini memiliki berat lebih atau kurang 200 kDa dan sebagian besar dalam sel induk dari sistem saraf pusat. Mereka diekspresikan selama perkembangan neuron.
Patologi terkait
Ada banyak penyakit pada manusia yang berhubungan dengan filamen menengah.
Pada beberapa jenis kanker seperti melanoma ganas atau karsinoma payudara, misalnya, ko-ekspresi filamen vimentin dan keratin menengah menyebabkan diferensiasi atau interkonversi sel epitel dan mesenkimal.
Telah ditunjukkan secara eksperimental bahwa fenomena ini meningkatkan aktivitas migrasi dan invasif sel kanker, yang memiliki implikasi penting dalam proses metastasis karakteristik kondisi ini.
Eriksson and Collaborators (2009) meninjau berbagai jenis penyakit dan hubungannya dengan mutasi spesifik pada gen yang terlibat dalam pembentukan enam jenis filamen perantara.
Penyakit yang berkaitan dengan mutasi gen pengkodean untuk dua jenis keratin adalah epidermolisis bullast, hiperkeratosis epidermolitik, distrofi kornea, keratodermias dan banyak lainnya.
Filamen menengah tipe III terlibat dalam banyak kardiomiopati dan penyakit otot yang berbeda yang terkait terutama dengan distrofi. Selain itu, mereka juga bertanggung jawab atas katarak dominan dan beberapa jenis sklerosis.
Banyak sindrom dan gangguan neurologis terkait dengan filamen tipe IV, seperti Parkinson Parkinson. Demikian pula, cacat genetik pada filamen tipe V dan VI bertanggung jawab untuk pengembangan penyakit autosom yang berbeda dan terkait dengan fungsi nukleus sel.
Contohnya adalah Sindrom Progeria Hutchinson-Gilford, Dystrophy Muscle-Dreifuss Emery, antara lain.
Referensi
- Anderton, b. H. (1981). Filamles Menengah: Keluarga Struktur Homolog. Jurnal Penelitian Otot dan Motilitas Sel, 2(2), 141-166.
- Eriksson, J. DAN., Pallari, h., Robert, d., Eriksson, J. DAN., Dechat, t., Senyum, b.,… Goldman, R. D. (2009). Memperkenalkan filamors menengah: dari penemuan ke penyakit. Jurnal Investigasi Klinis, 119(7), 1763-1771.
- Fuchs, e., & Weber, K. (1994). Filamles menengah: Struktur, dinamika, fungsi dan penyakit. Annu. Putaran. Biochem., 63, 345-382.
- Hendrix, m. J. C., Seftor, e. KE., Chu, dan. W., Trevor, k. T., & Seftor, r. DAN. B. (seribu sembilan ratus sembilan puluh enam). Peran kenyamanan menengah dalam migrasi, invasi dan metastasis. Ulasan Kanker dan Metastasis, limabelas(4), 507-525.
- Herrmann, h., & Aebi, u. (2004). Filame menengah: Struktur molekul, mekanisme perakitan, dan integrasi ke dalam perancah intraseluler yang berbeda. Tinjauan Tahunan Biokimia, 73(1), 749-789.
- Herrmann, h., & Aebi, u. (2016). Filamles menengah: Struktur dan perakitan. Perspektif Cold Spring Harbor dalam Biologi, 8, 1-22.
- McLean, i., & Lane, b. (sembilan belas sembilan puluh lima). Filamors menengah di Diew. Opini Saat Ini dalam Biologi Sel, 7(1), 118-125.
- Steinert, hlm., & Roop, D. (1988). Biologi molekuler dan seluler film menengah. Tinjauan Tahunan Biokimia, 57(1), 593-625.
- Steinert, hlm., Jones, J., & Goldman, R. (1984). Filames menengah. Jurnal Biologi Sel, 99(1), 1-6.
- « Karakteristik gastrotrik, habitat, reproduksi, makanan
- Karakteristik masjid lunak, taksonomi, habitat, penggunaan »