

Fitur dan fungsi kotak tata
- 1986
- 579
- Tommie Smith
Itu Kotak tata, Dalam biologi sel, ini adalah urutan konsensus DNA yang ditemukan di semua garis keturunan organisme hidup dan secara luas dipertahankan. Urutannya adalah 5'-tataa-3 'dan dapat mengikuti beberapa adenin berulang.
Lokasi kotak di atas (atau sungai di atas, karena biasanya disebut dalam literatur) dari awal transkripsi. Ini terletak di promotor gen, di mana persatuan dengan faktor transkripsi akan terjadi. Selain faktor -faktor ini, RNA polimerase II biasanya bergabung dengan kotak Tata.
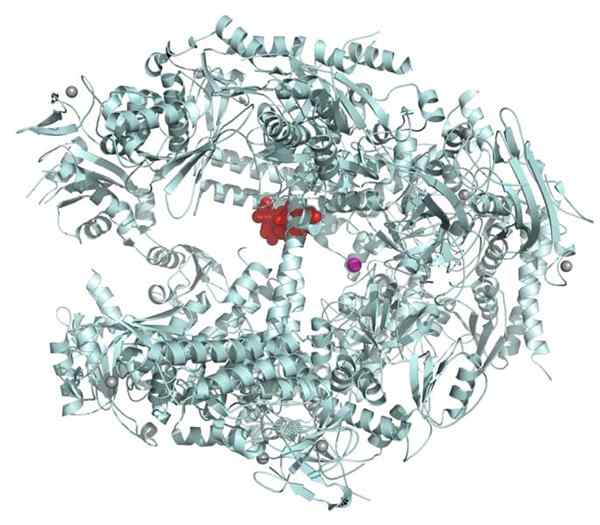
RNA polimerase II. Sumber: fvasconcellos 21:15, 14 November 2007 (UTC) [domain publik] Meskipun kotak tata adalah urutan utama promotor, ada gen yang tidak memilikinya.
[TOC]
Karakteristik
Awal sintesis RNA mensyaratkan bahwa RNA polimerase dipersatukan oleh sekuens DNA spesifik, yang disebut promotor. Tata Caja adalah urutan konsensus dari seorang promotor. Itu disebut kotak pribiew di prokariota, dan kotak-goldberg-hogness di eukariota.
Dengan demikian, kotak Tata adalah wilayah yang diawetkan dalam DNA. Urutan dari banyak daerah awal transkripsi DNA menunjukkan bahwa urutan konsensus, atau urutan umum, adalah (5ʾ) t*a*taat*(3ʾ). Posisi yang ditandai dengan tanda bintang memiliki homologi tinggi. Residu t terakhir selalu dalam promotor DAN. coli.
Lokasi kotak Tata di Procariota
Dengan konvensi, pasangan basa yang sesuai dengan awal sintesis molekul RNA diberi bilangan positif, dan pasangan basa yang mendahului awal RNA diberi bilangan negatif. Kotak Tata ada di wilayah -10.
Dapat melayani Anda: erythroblass: Apa itu erythropoiesis, patologi terkaitDi dalam DAN. coli, Wilayah promotor adalah antara posisi -70 dan +30. Di wilayah ini ada urutan konsensus kedua (5ʾ) t*tg*aca (3ʾ) di posisi -35. Demikian juga, posisi yang ditandai dengan tanda bintang memiliki homologi tinggi.
Lokasi kotak tata di eukariota
Dalam eukariota, daerah mempromosikan memiliki elemen sinyal yang berbeda untuk masing -masing RNA polimerase. Di dalam DAN. coli RNA polimerase tunggal mengidentifikasi elemen sinyal di wilayah promotor.
Selain itu, dalam eukariota daerah promosi lebih luas. Ada urutan yang berbeda, terletak di wilayah -30 dan -100, yang menetapkan kombinasi yang berbeda di berbagai promotor.
Dalam eukariota, ada banyak faktor transkripsi yang berinteraksi dengan promotor. Misalnya, faktor TFIID berikatan dengan urutan Tata. Di sisi lain, gen RNA ribosomal disusun dalam bentuk beberapa gen, satu diikuti oleh yang lain.
Variasi dalam urutan konsensus daerah -10 dan -35 mengubah penyatuan RNA polimerase ke wilayah promotor. Dengan demikian, mutasi basa tunggal menghasilkan penurunan kecepatan serikat RNA polimerase ke wilayah promotor.
Fungsi
Kertas transkripsi
Kotak Tata berpartisipasi dalam Union dan Inisiasi Transkripsi. Di dalam DAN. coli, Holoenzyme RNA polimerase terdiri dari lima subunit α2ββσ. Subunit σ bergabung dengan DNA rantai ganda dan bergerak mencari kotak Tata, yang merupakan sinyal yang ditunjukkan oleh awal gen.
Bagaimana transkripsi terjadi?
Subunit σ dari RNA polimerase memiliki konstanta yang sangat tinggi dari asosiasi dengan promotor (dalam urutan 10sebelas), yang menunjukkan spesifisitas pengakuan yang tinggi antara itu dan urutan kotak Pribiew.
Dapat melayani Anda: sel Langerhans: karakteristik, morfologi, fungsiRNA polimerase bergabung dengan promotor dan membentuk kompleks tertutup. Kemudian, itu membentuk kompleks terbuka yang ditandai dengan pembukaan lokal 10 pasangan dasar baling -baling DNA ganda. Pembukaan ini difasilitasi karena urutan kotak Pribiew kaya A-T.
Ketika DNA dibuka gulungan, tautan fosfodiéster pertama terbentuk dan argang RNA dimulai. Subunit σ dilepaskan dan RNA polimerase meninggalkan promotor. Molekul RNA polimerase lainnya dapat bergabung dengan promotor dan memulai transkripsi. Dengan cara ini gen dapat ditranskripsikan berkali -kali.
Dalam ragi, RNA polimerase II terdiri dari 12 subunit. Enzim ini memulai transkripsi yang mengenali dua jenis sekuens konsensus di ujung 5ʾ awal transkripsi, yaitu: konsensus Tata; Urutan konsensus CAAT.
Faktor transkripsi
RNA polimerase II membutuhkan protein, yang disebut faktor transkripsi TFII, untuk membentuk kompleks transkripsi aktif. Faktor -faktor ini cukup terpelihara di semua eukariota.
Faktor transkripsi adalah molekul sifat protein yang dapat bergabung dengan molekul DNA dan memiliki kemampuan untuk meningkatkan, mengurangi atau membatalkan produksi gen tertentu. Acara ini sangat penting untuk regulasi gen.
Pembentukan kompleks transkripsi dimulai dengan pengikatan protein TBP (“protein pengikat Tata”) dengan Tata Caja. Pada gilirannya, protein ini mengikat TFIIB, yang juga berikatan dengan DNA. Kompleks TBP-TFIIB bergabung dengan kompleks lain yang dibentuk oleh TFIIF dan RNA polimerase II. Dengan cara ini, TFIIF membantu RNA polimerase II untuk bergabung dengan promotor.
Dapat melayani Anda: lenticelaPada akhirnya, tfiie dan tfiih bergabung dan membuat kompleks tertutup. Tfiih adalah helosa dan mempromosikan pemisahan rantai DNA ganda, suatu proses yang dibutuhkan ATP. Ini terjadi di dekat awal sintesis RNA. Dengan cara ini, kompleks terbuka terbentuk.
Faktor transkripsi dan kanker
Protein p53 adalah faktor transkripsi, juga dikenal sebagai protein penekan tumor p53. Ini adalah produk dari kanker yang dominan. Sindrom Li-Francoi diproduksi oleh salinan gen bermutasi ini, yang menyebabkan penampilan karsinoma, leukemia dan tumor.
Diketahui bahwa p53 menghambat transkripsi beberapa gen dan mengaktifkan yang lain. Sebagai contoh, p53 mencegah transkripsi gen dengan promotor Tata melalui pembentukan kompleks yang dibentuk oleh p53, faktor transkripsi lainnya dan promotor Tata. Dengan demikian, p53 menjaga pertumbuhan sel tetap terkendali.
Referensi
- Bohinski, r. 1991. Biokimia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, hlm., Baltimore, d., Darnell, J. 2003. Biologi seluler dan molekuler. Editorial Pan -American, Buenos Aires.
- Teman-teman. 1994. P53: Sekilas tentang boneka di balik permainan bayangan. Sains, 265: 334.
- Devlin, t.M. 2000. Biokimia. Editorial Reverté, Barcelona.
- Voet, d., Voet, J. 2004. Biokimia. Jonh Wiley and Sons, New York.
- Nelson, d. L., Cox, m. M. 2008. Lehninger-prinsip biokimia. W.H. Freeman, New York.
- « Pelatihan, karakteristik, fungsi, penyakit
- Karakteristik Porphyromonas gingivalis, morfologi, siklus hidup »