

Struktur Sitosin, Fungsi, Properti, Sintesis
- 668
- 173
- Miss Marion Graham
Itu Sitosin Ini adalah basis nitrogen pirimidin, yang berfungsi untuk biosintesis cytidine-5'-monophosphate dan dexicitidine-5'-monophosphate. Senyawa ini masing -masing berfungsi untuk biosintesis, dari asam deoksiribonukleat (DNA) dan asam ribonukleat (RNA). DNA menyimpan informasi genetik dan RNA memiliki berbagai fungsi.
Pada makhluk hidup, sitosin tidak ditemukan secara bebas, tetapi biasanya ribonukleotida atau deoksiribonukleotida. Kedua jenis senyawa memiliki gugus fosfat, ribosa dan basa nitrogen.
Sumber: VESPRCOM [Domain Publik] Karbon 2 dari ribosa memiliki gugus oxhydrillo (-OH) dalam ribonukleotida, dan atom hidrogen (-H) dalam deoksiribonukleotida. Bergantung pada jumlah gugus fosfat yang ada, ada citidín-5'-monophosphate (CMP), citidín-5'-diphosphate (CDP) dan citidín-5'-triffosphate (CTP) (CTP).
Setara desoksigenasi disebut desoxicitidin-5'-monophosphate (DCMP), desoxicitidin-5'-diphosphate (DCDP), dan desoxicitidin-5'-triffosphate (DCTP).
Sitosin, dalam berbagai bentuknya, berpartisipasi dalam fungsi yang berbeda, seperti biosintesis DNA dan RNA, biosintesis glikoprotein dan regulasi ekspresi gen.
[TOC]
Struktur dan sifat
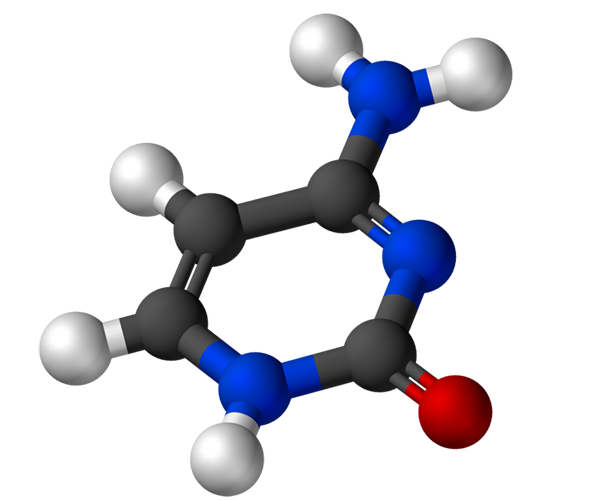
Sitosin, 4-amino-2-hydroxipirimidine, memiliki formula empiris C4H5N3Atau, yang berat molekulnya adalah 111,10 g/mol, dan dimurnikan sebagai bubuk putih.
Struktur sitosin adalah cincin heterosiklik, aromatik dan planar. Panjang gelombang absorbansi maksimum (ʎMax) Ini 260 nm. Suhu fusi sitosin melebihi 300 ºC.
Untuk membentuk nukleotida, sitosin mengikat secara kovalen, melalui nitrogen 1, dengan cara. Karbon 5 'diesterifikasi dengan gugus fosfat.
Biosintesis
Biosintesis nukleotida pirimidin memiliki jalur umum, yang terdiri dari enam langkah yang dikatalisis oleh enzim. Jalan dimulai dengan biosintesis karbamoil-fosfat. Dalam prokariota hanya ada satu enzim: karbamoil fosfat synthase. Ini bertanggung jawab untuk sintesis pirimidin dan glutamin. Pada eukariota, ada karbamoil fosfat sintase I dan II, yang masing -masing bertanggung jawab untuk biosintesis glutamin dan pirimidin.
Langkah kedua terdiri dari pembentukan N-karbamoilaspartato, dari karboil fosfat dan aspartat, reaksi yang dikatalisis oleh transpartat transcabamoilasa (atcasa).
Langkah ketiga adalah sintesis L-Dihydrotoate, yang menghasilkan penutupan cincin pirimidin. Langkah ini dikatalisasi oleh dihydrootase.
Langkah keempat adalah pembentukan orotato, yang merupakan reaksi redoks yang dikatalisis oleh dihydroorotate dehydrogenase.
Langkah kelima terdiri dari pembentukan goldylate (OMP) menggunakan fosforibosil pirofosfat (PRPP) sebagai substrat, dan transferase fosforibosil orotat sebagai katalis sebagai katalis.
Langkah keenam adalah pembentukan uridilaato (uridin-5'-monophosphate, emp.
Itu bisa melayani Anda: flora dan fauna dari hidalgoLangkah -langkah berikut terdiri dari fosforilasi UMP, dikatalisis oleh kinase, untuk membentuk UTP, dan transfer gugus amino dari glutamin ke UTP untuk membentuk CTP, reaksi yang dikatalisis oleh CTP sintetase.
Peraturan biosintesis
Pada mamalia, regulasi berlangsung di karbamoil fosfat sintase.
Carbamoil syntasa II diatur oleh umpan balik negatif. Regulatornya, UTP dan PRPP, masing -masing, adalah inhibitor dan aktivator enzim ini.
Dalam jaringan yang tidak hati, karbamoil fosfat sintase II adalah satu -satunya sumber karbamoil fosfat. Saat berada di hati, dalam kondisi amonia berlebih, karbamoil fosfat sintase I menghasilkan, di mitokondria, karbamoil fosfat, yang diangkut ke sitosol, dari mana ia memasuki jalur biosintesis pirimidines.
Titik regulasi lainnya adalah OMP-discarboxylase, yang diatur oleh penghambatan kompetitif. Produk reaksinya, UMP, bersaing dengan OMP untuk situs pengikatan di OMP-descarboxilasa.
Pirimidin, seperti sitosin, didaur ulang
Daur ulang pirimidin memiliki fungsi penggunaan kembali pirimidin tanpa perlu biosintesis novo, dan menghindari jalur degradatif. Reaksi daur ulang dikatalisis oleh fosforibosiltransferase pirimimidin. Reaksi umum adalah sebagai berikut:
Pirimidin + prpp -> nukleosida pirimidin 5 '-monofosfat + ppi
Pada vertebrata, fosforibosiltransferase pirimimidin ditemukan dalam eritrosit. Substrat pirimidin dari enzim ini adalah urasil, timina dan orotato. Sitosin secara tidak langsung didaur ulang dari uridin-5'-monophosphate.
Fungsi dalam biosintesis DNA
Selama replikasi DNA, informasi yang terkandung dalam DNA disalin dalam DNA melalui DNA polimerase.
Biosintesis RNA membutuhkan deoxynucleotides tryphosphate (DNTP), yaitu: dexitimid tryphosphate (dttp), dexicitidy triffosphate (DCTP), de -alxyxyadenin tryphyse (datp) dan dexiguanin tryfosphat. Reaksinya adalah:
(DNA)n limbah + DNTP -> (DNA)N+1 limbah + ppi
Hidrolisis anorganik pirofosfat (PPI) memberikan energi untuk biosintesis RNA.
Fungsi dalam stabilisasi struktur DNA
Dalam heliks DNUS ganda, purin, dari rantai, terkait dengan pirimidin, dari rantai yang berlawanan, oleh ikatan hidrogen. Dengan demikian, sitosin selalu terkait dengan guanin oleh tiga ikatan hidrogen: adenin terkait dengan timin oleh dua ikatan hidrogen.
Itu dapat melayani Anda: pentingnya biologiIkatan hidrogen rusak saat larutan DNA asli yang dimurnikan pada pH 7, mengalami suhu yang lebih besar dari 80 ºC. Ini menyebabkan baling -baling DNA ganda membentuk dua rantai terpisah. Proses ini dikenal sebagai denaturasi.
Suhu di mana 50% DNA didenaturasi dikenal sebagai suhu leleh (TM). Molekul DNA yang guanin dan sitosinnya.
Di atas adalah uji eksperimental bahwa jumlah ikatan hidrogen yang lebih besar menstabilkan molekul DNA asli.
Fungsi daerah kaya dalam sitosin dalam DNA
Baru -baru ini, ditemukan bahwa DNA nukleus sel manusia dapat mengadopsi struktur motif interkalasi (IM). Struktur ini diproduksi di daerah sitosin.
Struktur IM terdiri dari empat rantai DNA, tidak seperti DNA baling -baling ganda klasik yang memiliki dua rantai. Lebih khusus lagi, dua rantai dupleks paralel diselingi dalam orientasi antiparallela, dan dipertahankan bersama oleh sepasang sitosin hemiproton (C: C: C+).
Dalam genom manusia, struktur IM ditemukan di daerah seperti promotor dan telomer. Jumlah struktur IM lebih tinggi selama fase G1/S dari siklus sel, di mana transkripsi tinggi. Daerah -daerah ini adalah situs pengakuan protein yang terlibat dalam aktivasi mesin transkripsional.
Di sisi lain, di daerah yang kaya di pangkalan guanin (c) berturut -turut, DNA cenderung mengadopsi bentuk baling -baling A, dalam kondisi dehidrasi. Bentuk ini khas dari RNA dan pita campuran DNA-ARN ganda selama transkripsi dan replikasi, dan pada waktu-waktu tertentu ketika DNA terkait dengan protein.
Telah ditunjukkan bahwa daerah dengan basis sitosin berturut -turut membuat tambalan elektropositif di celah utama DNA. Oleh karena itu, diyakini bahwa daerah ini berikatan dengan protein, yang merupakan daerah genom tertentu dari kerapuhan genetik.
Fungsi dalam biosintesis RNA
Selama transkrip, informasi yang terkandung dalam DNA disalin dalam RNA melalui RNA PON POLYMERASE RNA. Biosintesis RNA membutuhkan nukleosida tryposphate (NTP), yaitu: tryphyse citidine (CTP), tryposphate uridine (UTP), adenine tryposphate (ATP) dan tryfosphate guanine (GTP). Reaksinya adalah:
Dapat melayani Anda: flora dan fauna Campeche: spesies representatif(RNA)n limbah + NTP -> (RNA)N+1 limbah + ppi
Hidrolisis anorganik pirofosfat (PPI) memberikan energi untuk biosintesis RNA.
Fungsi dalam biosintesis glikoprotein
Transfer sekuensial heksosis untuk membentuk oligosakarida, o-ligat ke protein, terjadi dari prekursor nukleotida.
Pada vertebrata, langkah terakhir dari biosintesis oligosakarida oigosakarida yang ligasi O terdiri dari penambahan dua limbah asam sinal (N-acetylneuramine) dari prekursor citidin-5'-monophosphate (CMP). Reaksi ini diproduksi dalam karung trans golgi.
Perawatan sitosin dan kemoterapi melawan kanker
Asam Tetrahydropholate (FH4) adalah sumber kelompok -CH3, Dan perlu untuk biosintesis DTMP dari dump. FH2 juga terbentuk. Pengurangan FH2 ke FH4 membutuhkan folat dan reduktase NADPH. Beberapa inhibitor folat reduktase, seperti aminopterin dan metretothrexate digunakan dalam perawatan kanker.
Metotrexan adalah penghambat kompetitif. Reduktase folat bergabung dengan afinitas 100 kali lebih banyak dengan inhibitor ini daripada substratnya. Aminapterine bertindak dengan cara yang sama.
Penghambatan folat reduktase secara tidak langsung menghambat biosintesis DTMP, dan oleh karena itu DCTP. Penghambatan langsung terjadi melalui inhibitor timidilate synthetase, yang mengkatalisasi DTMP dari dump. Inhibitor ini adalah 5-fluorouracil dan 5-fluoro-2-dioxiuridine.
Sebagai contoh, 5-fluoroacil bukanlah penghambat tetapi pertama-tama menjadi, rute daur ulang, dalam mofosfat d (FDUMP) deoksiurid, yang berikatan dengan synthesase timidilate dan menghambatnya.
Zat analog dengan glutamin, azaserin dan acivicin, menghambat amidotransferase glutamin. Azarine adalah salah satu zat yang ditemukan pertama kali yang bertindak sebagai tidak aktif bunuh diri.
Referensi
- Assi, h.KE., Garavia, m., González, c., Dan Damha, m.J. 2018. I-Motif DNA: Fitur Struktural dan Makna untuk Biologi Sel. Penelitian Asam Nuklei, 46: 8038-8056.
- Bohinski, r. 1991. Biokimia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokimia. Editorial Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, hlm., Baltimore, d., Darnell, J. 2003. Biologi seluler dan molekuler. Editorial Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Meksiko, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Lehninger-prinsip biokimia. W.H. Freeman, New York.
- Voet, d. dan voet, j. 2004. Biokimia. John Wiley and Sons, AS.