

Karakteristik, struktur, jenis, dan fungsi myosin
- 2721
- 840
- Jessie Harvey
Itu Myosin Ini adalah mesin molekuler, dari sifat protein, yang mampu bergerak pada filamen aktin dalam sitosol. Energi yang mendorong perpindahan myosin berasal dari hidrolisis ATP. Karena itu, myosin biasanya didefinisikan sebagai enzim mecanochemistry.
Dalam eukariota, myosin adalah protein yang sangat berlimpah. Ada berbagai jenis myosin, yang diberi kode oleh keluarga gen. Dalam ragi, 5 kelas dibedakan, sementara lusinan telah dijelaskan pada mamalia.
Sumber: David Richfield (Pengguna: SlashMe) Saat menggunakan gambar ini dalam karya eksternal, dapat dikutip sebagai berikut: Richfield, David (2014). "Galeri Medis David Richfield". Wikijournal of Medicine 1 (2). Doi: 10.15347/WJM/2014.009. ISSN 2002-4436. [CC BY-SA 3.0 (http: // createveCommons.Org/lisensi/by-sa/3.0/]] Myosin memiliki berbagai fungsi. Myosin I, di sebelah aktin, berpartisipasi dalam pergerakan keratosit.
Myosin II memberikan kekakuan pada membran plasma, berpartisipasi dalam sitokinesis dan kontraksi otot. Keduanya, myosinas I dan II, berkolaborasi dengan migrasi sel. Myosinas I dan V melakukan transportasi vesikel sepanjang filamen aktin.
[TOC]
Struktur
Dalam mikrograf elektronik, struktur isoform myosin khas memiliki tiga domain: kepala, leher dan ekor. Melalui hidrolisis dengan chimotripsin, segmen yang terdiri dari kepala dan leher, yang disebut heavy meromiosine (hmm), dan segmen ekor, yang disebut meromiosin cahaya (LMM) diperoleh.
Penguasaan kepala adalah ujung terminal-N dari rantai berat, dan domain ekor adalah ujung terminal-C dari rantai cahaya.
Kelas myosin dapat dibedakan dengan jumlah rantai polipeptik yang menyusunnya, dan kelas kelimpahan dan rantai cahaya bersatu di sekitar.
Myosin I memiliki rantai polipeptida, yang membentuk kepala dan ekornya tidak memiliki daerah helisidal alfa. Sementara myosinas i dan v memiliki dua rantai polipeptida, dan di sana.
Myosinas I dan V memiliki situs serikat untuk calmodulin, yang mengatur dan memperbaiki CA+2, Dalam rantai cahaya. Myosina yang saya perbaiki CA+2 Dalam rantai cahaya, tetapi sangat berbeda dari calmodulin.
Karakteristik
Pada tingkat mekanis, myosinas memiliki tiga karakteristik, yaitu:
- Kepala myosin adalah domain motor yang maju dengan langkah -langkah rahasia: penyatuan myosin menuju ke filamen aktin, kecenderungannya dan pemisahan selanjutnya menghasilkan pergerakan myosin. Proses ini bersifat siklik dan tergantung pada ATP.
- Perubahan Konformasi: Hidrolisis molekul ATP digabungkan pada setiap langkah molekul myosin, dengan cara amplifikasi dan tingkat transmisi. Ini menyiratkan perubahan konformasi myosin yang besar.
Dapat melayani Anda: simetri radialLevel pertama amplifikasi dihasilkan oleh hilangnya kelompok gamma-fosfat ATP, yang memungkinkan reorganisasi elemen struktural di situs ATP Union. Reorganisasi ini dikoordinasikan dengan perubahan struktural di situs Union to Actin.
Tingkat amplifikasi kedua menyiratkan komunikasi perubahan konformasi di situs aktif ke komponen struktural karboksil terminal.
- Directionality: Telah ditemukan bahwa milik saya memiliki polaritas, atau arah terbalik, menjelang akhir (+) dari filamen aktin. Kesimpulan ini berasal dari percobaan leveling filamen aktin, menggunakan mikroskop optik fluoresensi.
Fungsi
Myosin, di sebelah aktin, berpartisipasi dalam kontraksi otot, adhesi seluler, sitokinesis, memberikan kekakuan pada membran kortikal dan perpindahan beberapa vesikel, di antara fungsi -fungsi lainnya.
Cacat myosin dapat menghasilkan kondisi patologis. Misalnya, cacat pada myosinas I dan V terkait masing -masing, dengan miopati dari miosin dan gangguan pigmentasi (sindrom Griselli). Sedangkan gangguan pada isoform myosin vi menyebabkan kehilangan telinga.
Kontraksi otot
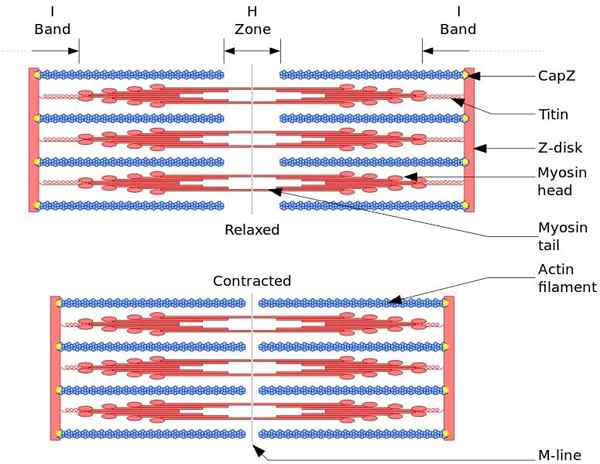
Unit fungsional dan struktural otot rangka adalah sarkomo. Selama kontraksi otot, panjang sarkomer mencapai 30% dari panjang aslinya.
Sarkomer terdiri dari filamen aktin yang tebal, myosin, dan tipis, yang disusun dengan cara yang kompleks. Secara umum, kepala myosin terletak di ujung distal filamen dan ekornya menuju pusat sarkomero, dan organisasi itu bipolar.
Untuk membuat kontraksi otot, kepala myosin, dari ujung yang berlawanan, harus pindah ke disk z atau ujung (+) filamen. Karena organisasi filamen tebal adalah bipolar, geser filamen tipis pada filamen tebal terjadi, digerakkan oleh ATP.
Gaya perpindahan terjadi karena ratusan kepala myosin, dari filamen tebal, berinteraksi dengan filamen tipis.
Sitosinisis
Selama mitosis, ketika mikrotubulus kutub gelendong dipisahkan, aktin dan myosin II membentuk cincin kontraktil dalam sel sel. Cincin ini mengontrak diameternya dan membagi sel menjadi dua bagian.
Pemberian kekakuan pada selaput kortikal
Dalam sel mutan yang kekurangan myosin II, membran plasma mudah berubah bentuk ketika gaya eksternal diterapkan. Ini terjadi karena myosin II memberikan kekuatan agregasi pada protein membran plasma.
Dapat melayani Anda: adaptasi morfologisAdhesi seluler
Dalam jaringan epitel, aktin kontraktil dan balok myosin II terletak di lingkungan membran plasma, dan membentuk sabuk melingkar yang mengelilingi permukaan sel seluler. Sabuk melingkar ini menentukan bentuk sel dan mempertahankan penyatuan di antara sel -sel.
Kontak antar sel terjadi dengan molekul sabuk melingkar ke molekul adhesi sel, menggunakan protein pengikat.
Perpindahan beberapa vesikel
Bukti eksperimental mengungkapkan bahwa myosin V melakukan transportasi membran dari peralatan Golgi ke pinggiran sel. Beberapa bukti adalah:
- Dalam sel jaringan saraf, dengan imunofluoresensi astrosit ditemukan bahwa myosin V terletak di sebelah Golgi.
- Dalam ragi, mutasi pada gen myosin V mengganggu sekresi protein dan, akibatnya, protein terakumulasi dalam sitosol.
- Myosin I isoform yang bertugas mengangkut vakuola ke membran sel. Menggunakan antibodi spesifik terhadap isoform myosin I, ditemukan bahwa isoform ini terletak di berbagai bagian sel.
Misalnya, ketika amuba hidup ditandai dengan antibodi terhadap myosin ic, pengangkutan vakuola ke membran ditangkap. Karena itu, vakuola mengembang dan sel meledak.
Penyakit terkait myosin
Myosinas dan kehilangan telinga
Ada banyak gen dan mutasi yang menghasilkan hilangnya telinga. Penyakit ini sering monogenetik .
Mutasi pada myosinas yang tidak konvensional, dengan satu atau dua kepala myosin, mempengaruhi fungsi telinga internal. Beberapa myosin myosin isaforms adalah myosin iiia, myosin viia dan myosin xva. Baru -baru ini, dua mutasi ditemukan dalam Myosin VI.
Mutasi pada myosin VI adalah c.897g> t dan p.926Q. Mutasi pertama mempengaruhi suatu daerah yang berinteraksi dengan lokasi yang aktif, yang disebut sakelar i. Homozigot untuk mutasi awal menunjukkan fenotipe, menyebabkan efek parah.
Mutasi kedua mempengaruhi daerah beban dengan beban, dalam baling -baling alfa di ekor myosin VI. Wilayah ini penting untuk dimerisasi proksimal mesin, dan mempengaruhi fungsi stereo-petak myosin VI.
Itu dapat melayani Anda: ilmu biologi tambahanMutasi lain adalah P.Asn207ser, yang menghasilkan mesin yang tidak dapat menghasilkan kekuatan. Ini karena ASN 207 adalah residu asam amino dari situs aktif, yang fungsinya adalah penyatuan dan hidrolisis ATP.
Mutasi hal.Arg657trp menghasilkan hilangnya fungsi myosin vi. Residu ARG terlibat dalam perubahan konformasi yang menempelkan hidrolisis ke gerakan myosin.
Myosin X dan Kanker
Myosina x (myo10) adalah myosin yang tidak konvensional yang diekspresikan di otak, endotelio dan banyak epitel. Myo10 dan tiga jenis proyeksi berbasis aktin (Philopoda, Invoicids dan Proyeksi Mirip dengan Philopodians) bekerja selama metastasis kanker.
Sel kanker invasif memiliki sejumlah besar philopoda dan mengekspresikan tingkat fascin yang tinggi. Protein ini melakukan intersects antara filamen aktin. Untuk melarikan diri dari tumor primer, penyerang terbentuk, kaya akan aktivitas proteolitik, yang mencerna matriks ekstraseluler di sekitarnya.
Setelah sel mencapai matriks ekstraseluler, proyeksi yang mirip dengan Philopoda membantu membubarkan dan menjajah. Tingkat tinggi MYO10 menunjukkan agresivitas dan metastasis yang tinggi pada kanker payudara.
Pembungkaman Myox menghasilkan hilangnya karakter metastasis sel, yang tidak dapat membentuk proyeksi berbasis aktin. Semua proyeksi ini memiliki adhesi berbasis komprehan, yang diangkut oleh MY1010 di dalam Philopodium.
Myox terlibat dalam pembentukan pusat. Tidak adanya Myox mendukung pembentukan lonjakan multipolar. Myox juga terlibat dalam pensinyalan dalam sel kanker. Misalnya, MYOX diaktifkan oleh 3,4,5, -inositol trifosfat (PIP3).
Referensi
- Alberts, b., Johnson, a., Lewis, J., dan kubis. 2007. Biologi molekul sel. Garland Science, New York.
- Brownstein, Z., Abu-Rayyan, a., Karfunkel-Don, d., Sirigu, s., Davido, b., Shohat, m., Frydman, m., Haussse, a., Kaanan, m., Avraham, k. 2014. Mutasi myosin novel untuk gangguan pendengaran herediter disukai oleh penangkapan genomik yang ditargetkan dan sekuensing paralel yang secara besar -besaran. Jurnal Genetika Manusia Eropa, 22: 768 -775.
- Courson, d.S. dan Cheney, R.DAN. 2015. Myosin-x dan penyakit. Penelitian Sel Eksperimental, 334: 10-15.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, hlm., Baltimore, d., Darnell, J. 2003. Biologi seluler dan molekuler. Editorial Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Meksiko, Sāo Paulo.
- Schliwa, m. dan woehlke, g. 2003. Motor molekuler. Alam, 422: 759 - 765.
- Untuk bernilai.D. 2003. Kotak alat molekuler untuk transportasi intelular. Sel, 112: 467-480.
- « Karakteristik, Struktur dan Fungsi Exonuclease
- Sejarah Bromo, Struktur, Konfigurasi Elektronik, Properti, Penggunaan »