

Struktur, fungsi, sifat, sintesis
- 2059
- 36
- Pete Lesch
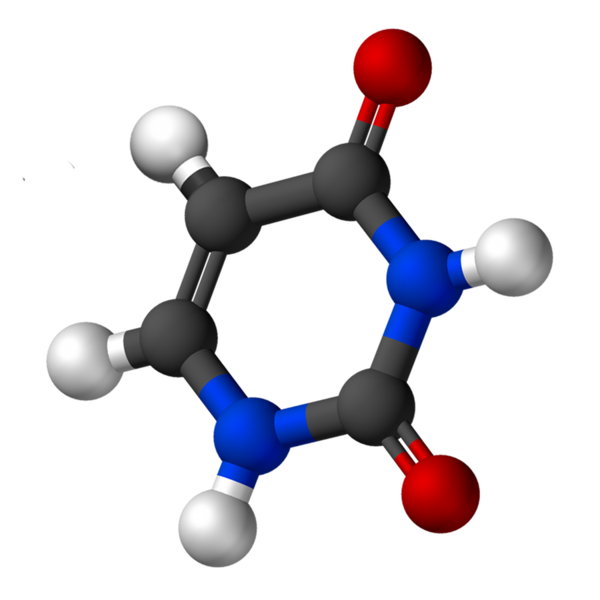
Dia Urasil Ini adalah basa nitrogen pirimidin, yang ditemukan dalam asam ribonukleat (RNA). Ini adalah salah satu karakteristik yang membedakan RNA dari asam deoksiribonukleat (DNA), karena yang terakhir memiliki timina, bukan urasil. Kedua zat, urasil dan timina, hanya berbeda bahwa yang kedua memiliki gugus metil.
Dari sudut pandang evolusi, telah diusulkan bahwa RNA adalah molekul pertama yang menyimpan informasi genetik dan bekerja sebagai katalis dalam sel, sebelum DNA dan enzim. Karena itu, diperkirakan bahwa urasil memiliki peran kunci dalam evolusi kehidupan.
Sumber: Kemikungen [domain publik] Pada makhluk hidup, urasil tidak ditemukan secara bebas, tetapi umumnya nukleotida monofosfat (UMP), difosfat (UDP) dan trphosphate (UTP). Nukleotida urasil ini memiliki fungsi yang berbeda, seperti biosintesis RNA dan glikogen, interkonversi isomerik gula dan regulasi sintamina sintase.
[TOC]
Struktur dan sifat
Uracil, disebut 2.4-dioxipiridine, memiliki formula empiris C4H4N2SALAH SATU2, yang berat molekulnya adalah 112,09 g/mol, dan dimurnikan sebagai bubuk putih.
Struktur uridin adalah cincin heterosiklik dengan empat atom karbon dan dua nitrogen, dengan ikatan alternatif ganda. Itu planar.
Ini memiliki kelarutan 50mg/ml, 25 ºC, dalam natrium hidroksida 1M, dan PKA antara 7.9 dan 8.2. Panjang gelombang di mana absorbansi maksimumnya terjadi (ʎMax) Ini antara 258 dan 260 nm.
Biosintesis
Ada jalur umum untuk biosintesis nukleotida pirimidin (urasil dan sitokin). Langkah pertama adalah biosintesis karbamoil fosfat dari co2 dan nh4+, yang dikatalisis oleh karbamoil sintetase fosfat.
Pirimidin dibangun dari karboil fosfat dan aspartat. Kedua zat bereaksi dan membentuk N-karbamoilaspartato, reaksi yang dikatalisis oleh transcabamoilasa aspartate (atcasa). Penutupan cincin pirimidin dapat didehidrasi oleh dihydrootase, dan menghasilkan L-dihidrotatoat.
L-Dihydrotoate teroksidasi dan diubah menjadi orotate; Akseptor elektron adalah NAD+. Ini adalah reaksi yang dikatalisis oleh dihydroorotate dehydrogenase. Langkah selanjutnya terdiri dari transfer gugus fosforibosyl, dari fosforibosil pirofosfat (PRPP), ke orotate. Formulir Gradidilato (WPO) dan Piroffosphat anorganik (PPI), dikatalisis oleh transferase fosforibosyl orotate.
Langkah terakhir terdiri dari dekarboksilasi cincin pirimidin kelompok (WPO). Bentuk uridilaato (uridin-5'-monophosphate, wasp), yang dikatalisis oleh dekarboksilase.
Kemudian, melalui partisipasi kinase, gugus fosfat ditransfer dari ATP ke UMP, membentuk UDP (uridin-5'-difosfat). Yang terakhir diulang, membentuk UTP (uridin-5'-triffosphate).
Dapat melayani Anda: cabang biologi dan apa yang mereka pelajariPeraturan biosintesis
Pada bakteri, regulasi biosintesis pirimidin terjadi melalui umpan balik negatif, pada level transcabamoilasa aspartate (ATCASA).
Enzim ini dihambat oleh CTP (citidin-5'-trophosphate), yang merupakan produk akhir dari jalur biosintesis pirimidin. ATCAS.
Pada hewan, regulasi biosintesis pirimidin terjadi melalui umpan balik negatif, pada tingkat dua enzim: 1) carbamoil sintase II fosfat, yang dihambat oleh UTP dan diaktifkan oleh ATP dan PRPP; dan 2) omp diskarboksilase, yang dihambat oleh produk reaksi yang dia sendiri katalis, UMP. Kecepatan biosintesis OMP bervariasi dengan ketersediaan PRPP.
Fungsi dalam biosintesis RNA
Uracil hadir di semua jenis RNA, seperti RNA messenger (MNA), transfer RNA (ARNT) dan RNA ribosom (RNA). Biosintesis molekul -molekul ini terjadi melalui proses yang disebut transkripsi.
Selama transkrip, informasi yang terkandung dalam DNA disalin dalam RNA melalui RNA PON POLYMERASE RNA. Proses terbalik, di mana informasi yang terkandung dalam RNA disalin dalam DNA, terjadi pada beberapa virus dan tanaman melalui transkriptase terbalik.
Biosintesis RNA membutuhkan nukleosida tryposphate (NTP), yaitu: tryphosphate uridine (UTP), tryposphate cytidine (CTP), adenine tryposphate adenine (ATP) dan tryphosphate guanine (GTP). Reaksinya adalah:
(RNA)n limbah + NTP -> (RNA)N+1 limbah + ppi
Hidrolisis anorganik pirofosfat (PPI) memberikan energi untuk biosintesis RNA.
Fungsi dalam biosintesis gula
Ester gula sangat umum dalam organisme hidup. Beberapa ester ini adalah difosfas ester nukleosid, seperti UDP-Sugar, yang sangat berlimpah di dalam sel. UDP-Sugar melibatkan biosintesis disakarida, oligosakarida dan polisakarida.
Pada tanaman, biosintesis sukrosa terjadi dengan dua cara: utama dan sekunder.
Rute utama terdiri dalam transfer D-glukosa dari UDP-D-glucosa ke D-Frucease untuk membentuk sukrosa dan UDP. Jalan sekunder termasuk dua langkah: dimulai dengan UDP-D-glukosa dan fruktosa-6-fosfat dan memuncak dengan pembentukan sukrosa dan fosfat.
Di kelenjar susu, biosintesis laktosa terjadi dari UDP-D-galaktosa dan glukosa.
Dapat melayani Anda: cabang biokimiaPada tanaman, biosintesis selulosa dilakukan dengan kondensasi kontinu limbah beta-D-glikosil, dari UDP-glukosa ke ujung yang tidak mengurangi rantai poliglukosa yang tumbuh. Demikian pula, biosintesis amilosa dan amilopektin membutuhkan UDP-glukosa sebagai substrat donor glukosa ke rantai yang tumbuh.
Pada hewan, baik UDP-glukosa dan ADP-glukosa digunakan untuk biosintesis glikogen. Demikian pula, biosintesis chondroitine sulfate membutuhkan UDP-xylose, UDP-galaktosa dan UDP-glucuronato.
Fungsi dalam Interkonversi Isomerik Gula
Konversi galaktosa menjadi perantara glikolisis terjadi melalui jalan leloir. Salah satu langkah rute ini dikatalisis oleh enzim UDP-galactose-4-epimerase, yang memfasilitasi interkonversi UDP-galaktosa menjadi UDP-glukosa.
Fungsi dalam biosintesis glikoprotein
Selama biosintesis glikoprotein, protein melewati cis, medium dan karung trans peralatan Golgi.
Masing -masing tas ini memiliki satu set enzim yang memproses glikoprotein. Monomer gula, seperti glukosa dan galaktosa.
Nukleotida-heksosa diangkut ke tangki golgi dengan antiporte. UDP-galactose (UDP-gal) dan UDP-N-acetylgalactosamina (UDP-Galnac) masukkan tangki dari sitosol dengan cara pertukaran dengan UMP.
Di tangki golgi, fosfatase menghidrolisis gugus fosfat dari bentuk UDP dan UMP dan PI. UDP berasal dari reaksi yang dikatalisis oleh galactosyltransferase dan N-acetylgalactosamiltransferase. Umpan yang dibentuk oleh fosfatase berfungsi untuk bertukar nukleotida-heksosa.
Fungsi dalam regulasi sintase
Mekanisme untuk mengatur glutamin sintase adalah modifikasi kovalen, yang terdiri dari adenilasi, yang tidak aktif, dan mengapit, yang aktif. Modifikasi kovalen ini reversibel dan dikatalisis oleh adenyltransferase.
Aktivitas adenyltransferase dimodulasi oleh penyatuan protein PII, yang diatur oleh modifikasi kovalen, uridinilasi.
Baik uridililation dan offset dilakukan oleh uridilransferase. Dalam enzim ini, aktivitas uridilasi disebabkan oleh glutamin dan fosfat, dan diaktifkan oleh penyatuan alfa-zotoglutarat dan ATP ke PII.
Fungsi dalam Edisi RNA
Beberapa RNM diedit sebelum terjemahan. Dalam beberapa organisme eukariotik, seperti Trypanosoma brucei, Ada edisi RNA dari transkrip gen subunit II dari sitokrom oksidase. Ini terjadi dengan memasukkan limbah urasil, reaksi yang dikatalisis oleh terminal uridiltransferase.
Dapat melayani Anda: curl: komposisi, bagian, kepentinganPanduan RNA, pelengkap produk yang diedit, bertindak sebagai sedang untuk proses pengeditan. Pasangan basa terbentuk antara transkrip awal dan panduan RNA menyiratkan pasangan basa g = u yang bukan watson-record dan umum di RNA.
Biosintesis UDP-glukosa
Dalam kondisi fisiologis, biosintesis glikogen dari glukosa-1-fosfat secara termodinamik tidak mungkin (ΔG positif). Karena itu, sebelum biosintesis, aktivasi glukosa-1-fosfat (G1P) terjadi. G1P dan UTP ini menggabungkan reaksi untuk membentuk glukosa difosfat uridin (UDP-glukosa atau UDPG).
Reaksi dikatalisis oleh pirofosforlase dari UDP-glukosa, dan adalah sebagai berikut:
G1p + utp -> udp -glucosa + 2pi.
Variasi energi bebas Gibbs dalam langkah ini besar dan negatif (-33,5 kJ/mol). Selama reaksi oksigen, G1P menyerang atom alfa alfa fosfor alfao dari UDP-glukosa dan anorganik pirofosfat (PPI). Kemudian, PPI dihidrolisis oleh pirofosfat anorganik, yang energi hidrolisisnya adalah yang menggerakkan reaksi umum.
UDP-glukosa adalah zat "berenergi tinggi". Ini memungkinkan untuk membentuk ikatan glikosida antara residu glukosa dan peningkatan rantai polisakarida. Prinsip energi yang sama ini berlaku untuk reaksi di mana UDP-Sugar berpartisipasi, seperti biosintesis disakarida, oligosakarida dan glikoprotein.
Uracil DNA glycosilasa
Ada lesi DNA yang terjadi secara spontan. Salah satu lesi itu adalah sprontane. Dalam hal ini, perbaikan terjadi karena basis DNA yang dimodifikasi dengan menggunakan enzim yang disebut Uracil DNA glycosilasa.
Enzim Uracil DNA glikosilasa menghilangkan sitokin yang rusak (urasil), menghasilkan residu deoksiribosa yang tidak memiliki basis nitrogen, yang disebut situs AP (situs apurinic-apirimidinic).
Kemudian, enzim AP endonuklease membuat potongan kerangka fosfodiester dari situs AP, menghilangkan residu gula-fosfat. DNA Polymerase I mengembalikan untaian yang rusak.
Referensi
- Bohinski, r. 1991. Biokimia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokimia. Editorial Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, hlm., Baltimore, d., Darnell, J. 2003. Biologi seluler dan molekuler. Editorial Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Meksiko, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Lehninger-prinsip biokimia. W.H. Freeman, New York.
- Voet, d. dan voet, j. 2004. Biokimia. John Wiley and Sons, AS.