

Karakteristik, fungsi, komponen pori nuklir
- 1459
- 175
- Ernesto Mueller
Dia pori nuklir (Dari bahasa Yunani, pori -pori = Langkah atau Transit) adalah "pintu" nukleus yang memungkinkan sebagian besar transportasi antara nukleoplasma dan sitoplasma. Pori nuklir bergabung dengan membran internal dan eksternal nukleus untuk membentuk saluran, yang berfungsi untuk pengangkutan protein dan RNA.
Kata poro tidak mencerminkan kompleksitas struktur yang dimaksud. Karena itu, lebih baik merujuk pada kompleks pori nuklir (CPN), daripada pori nuklir. CPN dapat mengalami perubahan dalam strukturnya selama transportasi atau keadaan siklus sel.
Baru -baru ini, telah ditemukan bahwa nukleoporin, protein yang membentuk CPN, memiliki peran penting dalam mengatur ekspresi gen. Dengan demikian, ketika mutasi terjadi yang mempengaruhi fungsi nukleoporin, patologi diproduksi pada manusia, seperti autoimun, kardiomiopati, infeksi virus dan kanker.
[TOC]
Karakteristik
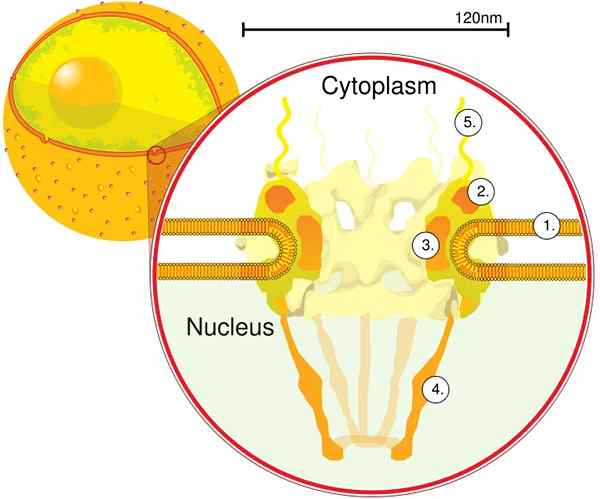
Melalui elektron tomografi ditentukan bahwa CPN memiliki ketebalan 50 nm, diameter eksternal antara 80 dan 120 nm, dan diameter internal 40 nm. Molekul besar seperti subunit besar ribosom (PM 1590 kDa) dapat diekspor di luar nukleus melalui CPN. Diperkirakan ada antara 2000 dan 4000 CPN per nukleus.
Berat molekul CPN individual kira -kira antara 120 dan 125 mda (1 mda = 106 Da) dalam vertebrata. Sebaliknya, CPN lebih kecil dalam ragi, di mana ia memiliki sekitar 60 mda. Meskipun ukuran CPN yang sangat besar, nukleoporin sangat terpelihara di semua eukariota.
Translokalisasi melalui CPN adalah proses yang cepat, yang kecepatannya adalah 1000 translokasi/kedua. Namun, CPN tidak menentukan arah aliran transportasi itu sendiri.
Ini tergantung pada gradien rangtp, yang lebih besar di nukleus daripada di sitosol. Gradien ini dipertahankan oleh faktor penukar guanin.
Selama siklus sel, CPN mengalami siklus perakitan dan perpecahan. Perakitan terjadi di antarmuka dan segera setelah mitosis.
Fungsi
Asam ribonukleat (RNA nuklir kecil, RNA messenger, RNA transfer), protein dan ribonukleoprotein (RNP) harus secara aktif diangkut melalui CPN. Ini berarti bahwa energi hidrolisis ATP dan GTP diperlukan. Setiap molekul diangkut dengan cara tertentu.
Secara umum, molekul RNA dikemas dengan kompleks RNP yang membentuk protein, diekspor dengan cara ini. Sebaliknya, protein yang secara aktif diangkut ke nukleus harus memiliki tanda urutan lokasi dalam nukleus (SLN), pemilik limbah asam amino dengan beban positif (misalnya, KKKRK).
Dapat melayani Anda: mesosomeProtein yang diekspor ke nukleus harus memiliki sinyal ekspor ke nukleus (NES) yang kaya asam amino leusin.
Selain memfasilitasi transportasi antara nukleus dan sitoplasma, CPN terlibat dalam organisasi kromatin, regulasi ekspresi gen dan perbaikan DNA. Nukleoporin (NUPS) mempromosikan aktivasi atau represi transkripsi, terlepas dari keadaan proliferasi sel.
Dalam ragi, nyaring ditemukan di CNP pembungkus nuklir. Di metazoaries mereka ada di dalam. Mereka melakukan fungsi yang sama di semua eukariota.
Impor zat
Melalui CPN ada penyebaran pasif molekul kecil di kedua arah dan transportasi aktif, impor protein, ekspor RNA dan ribonukleoprotein (RNP), dan antar -jemput molekul dua arah. Yang terakhir termasuk RNA, RNP dan protein yang terlibat dalam pensinyalan, biogenesis dan penggantian.
Impor protein dalam nukleus terjadi dalam dua langkah: 1) pengikatan protein ke sisi sitoplasma CPN; 2) Translocalization Dependent ATP melalui CPN. Proses ini membutuhkan hidrolisis ATP dan pertukaran GTP/PDB antara nukleus dan sitoplasma.
Menurut model transportasi, kompleks penerima-protein bergerak di sepanjang saluran melalui Union, disosiasi dan penyatuan lagi ke urutan FG yang berulang dari nukleoporin. Dengan cara ini, kompleks bergerak dari satu nukleoporin ke yang lain di dalam CPN.
Ekspor zat
Mirip dengan impor. Berlari GTPase memaksakan arah pada transportasi melalui CNP. RAN adalah sakelar molekuler dengan dua keadaan konformasi, tergantung pada apakah itu terkait dengan PDB atau GTP.
Dua protein pengatur spesifik memicu konversi antara kedua keadaan: 1) sitosolik GTPase Activating Protein (GAP), yang menghasilkan hidrolisis GTP dan dengan demikian mengubah RAN-GTP menjadi RAN-GDP; dan 2) Faktor Pertukaran Guanina Nuklir (GEF), yang mempromosikan pertukaran PDB oleh GTP dan mengkonversi RAN-GDP menjadi RAN-GTP.
Sitosol terutama mengandung ran-gdp. Nukleus terutama mengandung ran-gtp. Gradien dari dua bentuk konformasi RAN ini mengarahkan transportasi di alamat yang sesuai.
Impor penerima, bersama dengan posisi, difasilitasi oleh Union untuk pengulangan-FG. Jika Anda mencapai sisi nuklir CNP, Ran-GTP bergabung dengan penerima untuk melepaskan posisi Anda. Dengan demikian, Ran-GTP menciptakan arah proses impor.
Ekspor nuklir serupa. Namun, Ran-GTP dalam nukleus mempromosikan penyatuan posisi ke penerima ekspor. Ketika penerima ekspor bergerak melalui pori menuju sitosol, ia memenuhi ran-gap, yang menginduksi hidrolisis GTP ke PDB. Akhirnya, penerima dilepaskan dari posisinya dan Ran-GDP di sitosol.
Dapat melayani Anda: leukosit polimorfonuklearTransportasi RNA
Ekspor beberapa jenis RNA mirip dengan ekspor protein. Misalnya, ARNT dan RNAN (nuklir kecil) menggunakan gradien rangtp dan diangkut melalui CPN dengan cara. Ekspor ribosom matang juga tergantung pada gradien rangtp.
MRNA diekspor dengan cara yang sangat berbeda dari protein dan RNA lainnya. Untuk ekspornya, mRNA membentuk kompleks RNP Messenger (RNPM), di mana molekul RNM dikelilingi oleh ratusan molekul protein. Protein ini memiliki pemrosesan, Capping, Empalme dan Polyadentilly of MNA.
Sel harus dapat membedakan antara RNPM dengan RNM dan RNPM dewasa dengan mRNA yang belum matang. MRNA, yang membentuk kompleks RPNM, dapat mengadopsi topologi yang perlu direnovasi untuk transportasi. Sebelum RNPM memasuki CPN, langkah kontrol yang dilakukan oleh tramp dan kompleks protein eksosom terjadi.
Ketika RNPM dewasa dirakit, RPNM diangkut melalui saluran dengan menggunakan penerima transportasi (NXF1-NXT1). Penerima ini membutuhkan hidrolisis ATP (bukan gradien rangtp) untuk menetapkan renovasi arah RNPM, yang akan mencapai sitoplasma.
Kompleks pori nuklir dan kontrol ekspresi gen
Beberapa penelitian menunjukkan bahwa komponen CPN dapat mempengaruhi regulasi ekspresi genetik melalui kontrol struktur kromatin dan aksesibilitasnya ke faktor transkripsi.
Dalam eukariota evolusi yang lebih baru, heterokromatin lebih disukai terletak di pinggiran nukleus. Wilayah ini terganggu oleh saluran euchromatin, yang dipertahankan oleh keranjang nuklir CPN. Asosiasi keranjang nuklir dengan euchromatin terkait dengan transkripsi gen.
Sebagai contoh, aktivasi transkripsi pada tingkat CPN menyiratkan interaksi komponen keranjang nuklir dengan protein seperti histone acetyltransferase saga dan faktor ekspor RNA.
Dengan demikian, keranjang nuklir adalah platform untuk banyak gen pemeliharaan (Pembenahan) sangat ditranskripsi dan gen yang sangat disebabkan oleh perubahan kondisi lingkungan.
Kompleks pori dan virologi nuklir
Infeksi sel Eukaryot virus tergantung pada CPN. Dalam setiap kasus infeksi virus, keberhasilannya tergantung pada DNA, RNA atau RPN yang melewati CPN untuk mencapai tujuan utamanya, yang merupakan replikasi virus.
Dapat melayani Anda: inklusi sitoplasma: apa itu, karakteristik, fungsiAPE Virus 40 (SV40) telah menjadi salah satu model yang paling banyak dipelajari untuk menyelidiki fungsi CPN dalam translokalisasi dalam nukleus. Ini karena SV40 memiliki genom kecil (5.000 pangkalan).
Telah ditunjukkan bahwa pengangkutan DNA virus difasilitasi protein protein virus, yang melindungi virus sampai nukleus mencapai.
Komponen
CPN tertanam di dalam amplop nuklir dan terdiri dari antara sekitar 500 dan 1000 NUPTS. Protein ini diatur dalam subkompleks atau modul struktural, yang berinteraksi satu sama lain.
Modul pertama adalah komponen sentral, atau cincin, di dalam pori dalam bentuk jam pasir, yang dibatasi oleh cincin berdiameter 120 nm lainnya di kedua sisi, intranuklear dan sitoplasma. Modul kedua adalah cincin nukleus dan sitoplasma (masing -masing dengan diameter 120 nm) yang terletak di sekitar komponen berbentuk pasir.
Modul ketiga adalah delapan filamen yang diproyeksikan dari cincin 120 nm di dalam nukleoplasma dan membentuk struktur berbentuk keranjang. Modul keempat terdiri dari filamen yang diproyeksikan ke sisi sitoplasma.
Kompleks berbentuk Y, yang terdiri dari enam protein NUPS dan SEH 1 dan SEC 13, adalah kompleks CNP terbesar dan berkarakter terbaik. Kompleks ini adalah unit penting yang merupakan bagian dari perancah CPN.
Meskipun kesamaan rendah antara urutan NUPS, perancah CPN sangat diawetkan di semua eukariota.
Referensi
- Beck, m., Terluka, e. 2016. Kompleks pori nuklir: Memahami fungsinya melalui wawasan struktural. Ulasan Alam, Biologi Sel Molekul, DOI: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Protein pori nuklir dan kontrol fungsi genom. Gen dan Pengembangan, 29, 337-349.
- Kabachinski, g., Schwartz, t.ATAU. 2015. Kompleks pori nuklir - struktur dan fungsi sekilas. Jurnal Ilmu Sel, 128, 423-429.
- Knocknhauer, k.DAN., Schwartz, t.ATAU. 2016. Kompleks pori nuklir sebagai gerbang yang fleksibel dan dinamis. Sel, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.W. 2014. Nuklir multifungsi dengan kompleks: Platform untuk mengendalikan ekspresi gen. Opini saat ini tentang biologi sel, doi: 10.1016/j.Ceb.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Publikasi: Kompleks Pori Nuklir - Tinjauan Komprehensif Struktur dan Fungsi. Jurnal Internasional Kedokteran Akademik, 3, 51-59.
- Tran, e.J., Wente, s.R. 2006. Nuklir dinamis oleh kompleks: Life on the Edge. Sel, 125, 1041-1053.